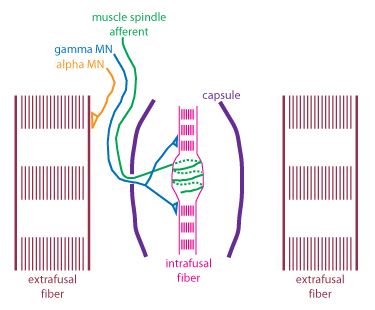
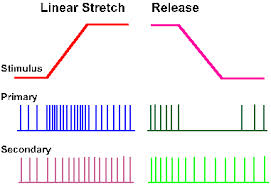
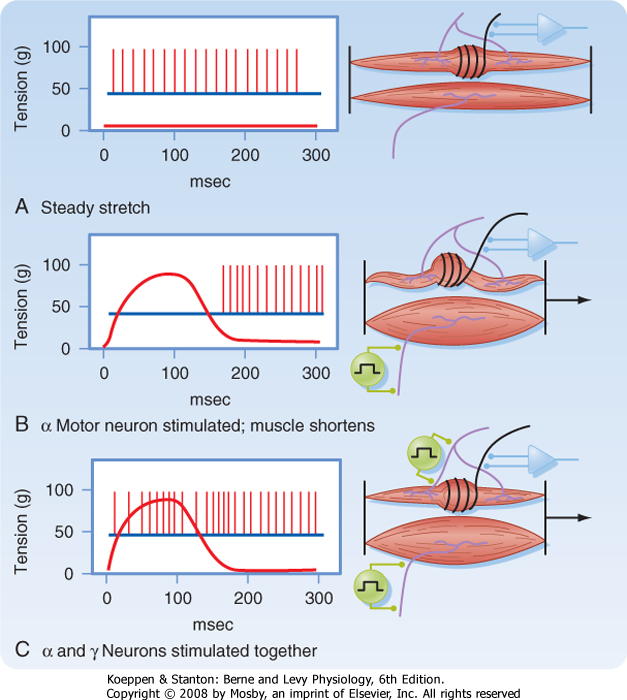
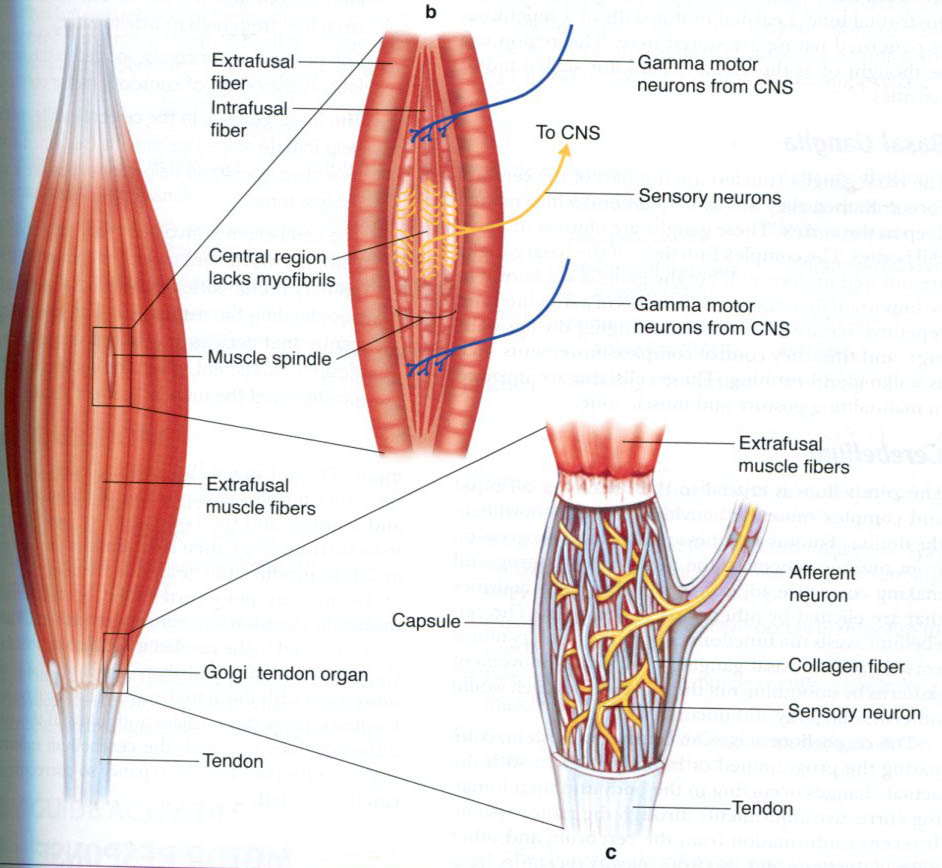
Sensory nerve endings can be classified according to morphology (Encapsulated vs Free nerve endings) or by their functional properties. Sensory receptors transduce tactile stimuli and signal the intensity and temporal changes in the forces applied to the skin by passing trains of action potentials to the spinal cord. Each single afferent axon provides information about events in the area of skin it innervates (the receptive field) and the position and direction of movement of a tactile stimulus are computed from the patterns of activity in many axons, as the stimulus moves from the receptive field of one neurone to another. There are several different types of nerve ending in the skin which each respond to different, specific aspects of the stimulus and there is a good correlation between the function and structure of these nerve endings. In skeletal muscle, there are also specific receptors that monitor the changes in length and tension of individual muscles, and there is also a good correlation between the function and structure of these nerve endings. |
| Structure of Sensory Nerve Endings | Location of Nerve Endings |
|
Adequate Stimulus Electrical recordings of action potential trains in single afferent nerve fibres ('single units') show that the information carried by each sensory axon relates to a specific type of natural stimulus ('modality' - mechanical, thermal, etc); their transducers are fairly specific for a particular form of natural stimulation. The 'adequate stimulus' is the specific natural stimulus (mechanical, thermal or noxious, etc) to which the transducers are most sensitive. The adequate stimulus can be used to classify sensory receptors. Most sensory endiings respond to one main type of stimulus, such as light touch, stretch, or temperature. Thus for the sensation of touch, there are around four different types of sensory neurones responding to:
Others respond to hair movement, and the hair receptors are most highly developed in the vibrissae of animals such as cats. Cutaneous receptors are classified into three main groups: Mechanoreceptors respond to mechanical stress or strain, or vibration. Thermoreceptors respond to the normal range of temperatures in the skin. Nociceptors respond to damaging stimuli such as painful mechanical chemical or thermal stimuli. The skin proteins begin to denature around 45 degees centigrade, and heat nociceptors respond to temperatures above 45 degrees C. The threshold stimulus is the intensity of the natural stimulus that causes the afferent endings to start producing action potentials. So the temperature threshold of thermoreceotprs is less than that of nociceptors. Also the threshold of touch afferents is less than that of the mechanical nociceptors that only respond to damaging stimuli. The receptive field is the area of skin that is innervated by a single sensory neurone. Some receptive fields are small, and a few may be described as punctate. But many axons innervate a moderately large area of skin, and the receptive fields of single axons overlap with each other; the exact location of a stimulus is worked out by the CNS as a result of comparisons of the spatial distribution of activity in a large number of axons. Overlapping receptive fields in the skin can also be demonstrated when a nerve is cut. The area of skin innervated by a single nerve trunk does not become insensitive following section of one dorsal root, because adjacent roots also innervate that area. Spatial discrimination may change, but the fact remains that the areas of skin innervated by single axons or nerve trunks overlap with others. |
Adaptation Rate of adaptation
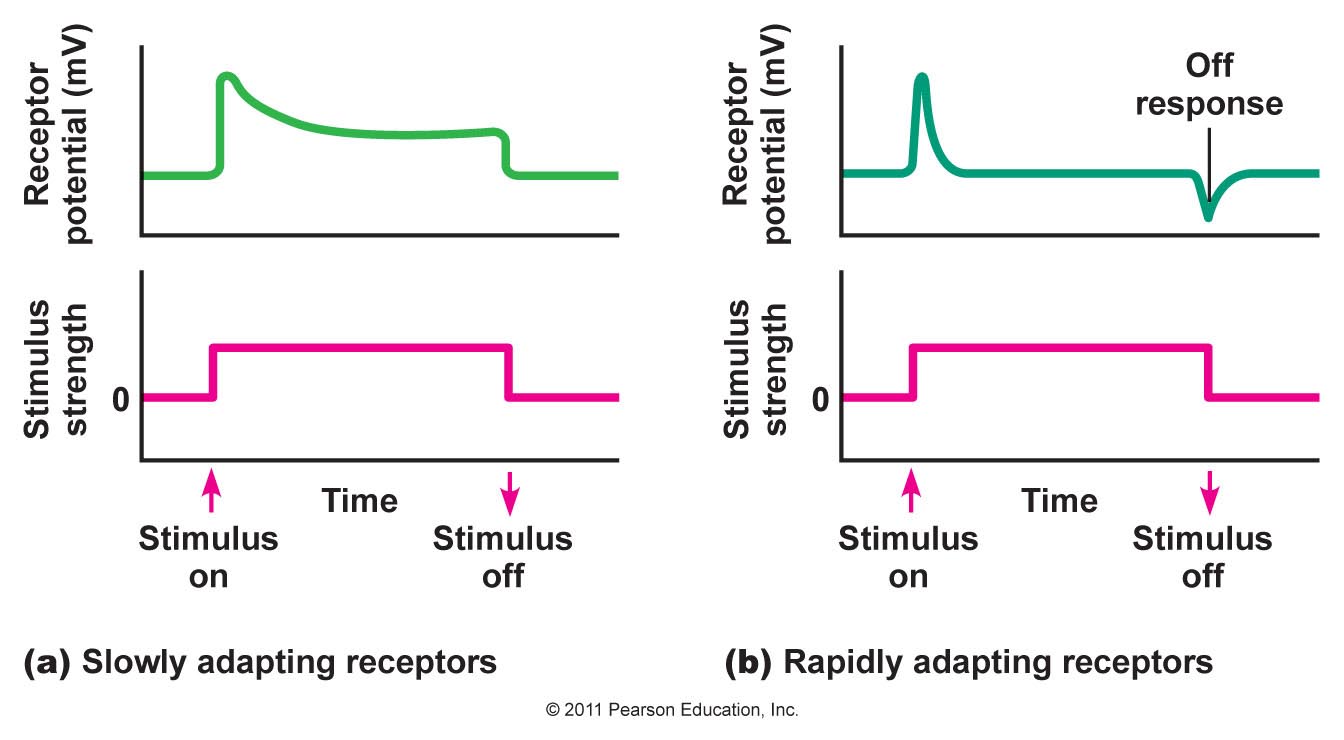
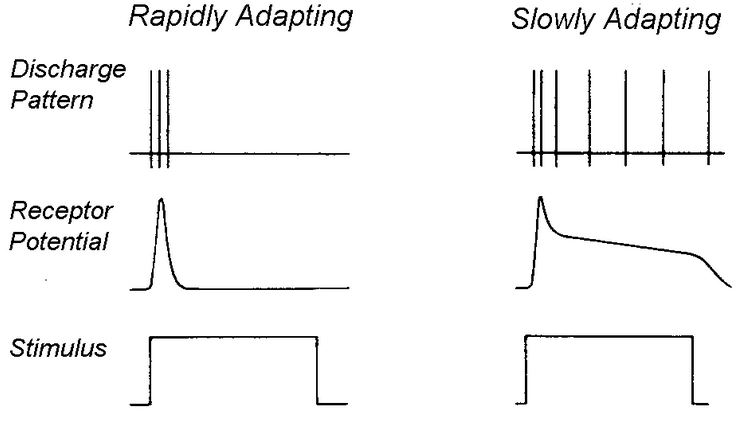
The receptor (generator) potential is the depolarising potential produced by the transducers at sensory nerve endings that initiates trains of action potentials in the afferent axon. It can be seen that in phasic (rapidly adapting) receptors the generator potential lasts only for the period during which the stimulus is applied. It can be seen from the diagram that many slowly adapting receptors also show some phasic activity at the time the stimulus is applied. Whereas in slowly adapting (tonic) receptors the generator potential is maintained throughout the natural stimulus. Some receptors monitor events inside the body : interoceptors include baroreceptors, chemoreceptors and osmoreceptors that sense arterial pressure, arterial blood gases or the osmolality of blood. This information is used to regulate the functions of internal organs, but does not enter consciousness. Others, in muscle tendons and joints, are proprioceptors, responsible for the sense of kinaesthesia- the sense of position of the limbs. |
Frequency Code The intensity of a natural stimulus is signalled by the frequency of action potentials generated during the tonic phase of their discharge. In rapidly adapting receptors, the rate of discharge of action potentials is related to the change in the generator potential, which is dependent on the rate of application of the stimulus. In slowly adapting receptors, the generator potential is maintained and that causes a train of impulses to occur in which the rate of discharge is related to the depolarisation. Most tonic, slowly adapting receptors, show some phasic activity related to the speed of onset of the stimulus. |
|
Axons may be classified according to their diameter or their conduction velocity: for myelinated axons, there is a relationship between the two, and as a rule of thumb, the conduction velocity in m/sec is roughly the six times the axonal diameter. The two forms of classification use different symbols: Groups A, B and C based on diameter, and Groups I, II, III and IV, based on conduction velocity. The former has been used in studies of skin nerves, and the latter in studies of muscle nerve afferents. |
|
Classification by diameter : the A, B, C system A fibres have large diameters, and C fibres are amall unmyelinated axons.
A fibres have diameters between 2 (A delta) and 20 A alpha Examples are ;
|
Classification by Conduction Velocity: the I, II, II, IV system Myelinated fibres in Muscle afferent nerves are divided into four goups depending on conduction velocity
Group I have higher conduction velocities than the other groups, with Group IV being the slowest. Examples are:
|