|
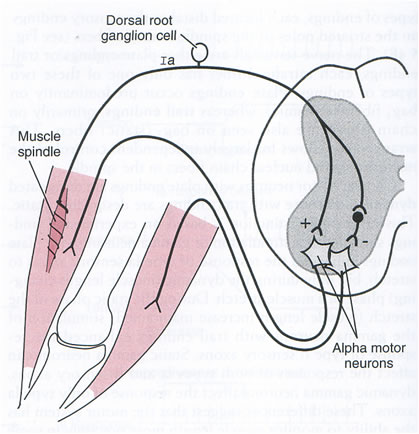
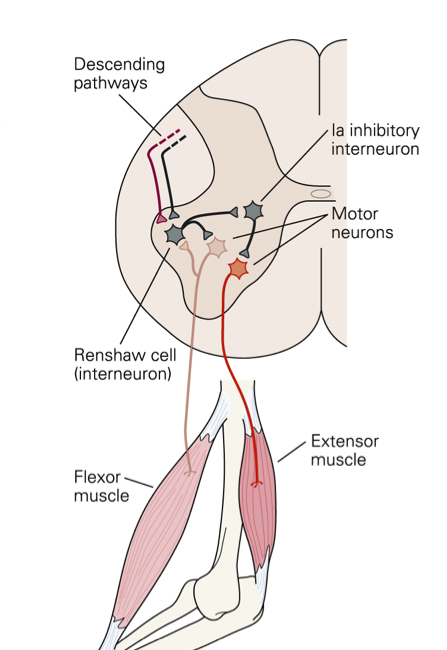
All reflexes have an afferent pathway that uses a specific type of sensory receptor, at least one synapse in the pathway, and an efferent pathway that connects to a muscle or gland. The site of the synapse is generally within the spinal cord, although the enteric nervous system also participates in reflex activity confined to the gut wall. Synaptic transmission is unidirectional – from pre-synaptic ending on to post-synaptic membrane, because the neurotransmitter is released from the pre-synaptic ending and acts on the receptors in the post-synaptic membrane. Some reflexes are monosynaptic, such as the stretch reflex. Other reflex pathways are polysynaptic, and include interneurones that connect the sensory and effector neurones. |
|
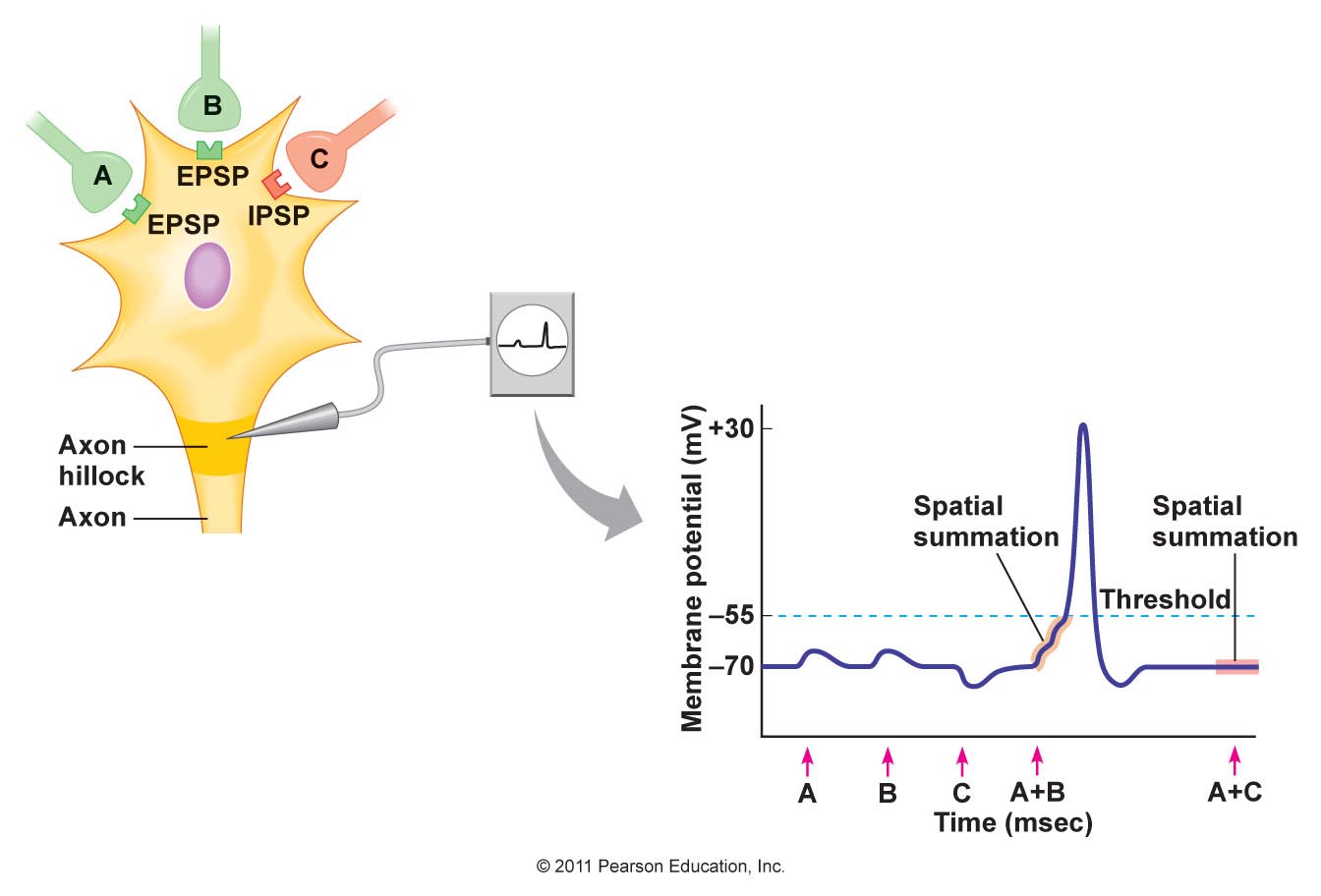
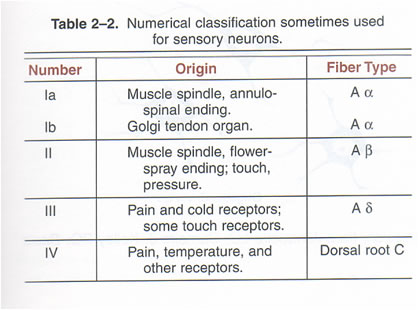
The timing of potentials recorded in the spinal cord The classification of nerve fibres according to conduction velocity is important in identifying the precise time of events in the reflex pathway. Electrical stimuli are applied to afferent nerves, and the stimulus intensity adjusted to recruit axons of different conduction veloicities. The arrival of these action potentials at the dorsal root can be monitored to be sure whcih groups of afferents are being stimulated: action potentials in Ia fibres arrive there before those in Ib axons, and the Group II potentials arrive later still. The timing of potentials recorded by microelectrodes in spinal cord neurones also depends on the number of synapses in the pathway, as each synapse is associated with a synaptic delay of 0.2-0.5 msec. Consequently, events in neurones involved in the monosynaptic reflex occur before those in neurones of the flexor reflex, etc, and the duration of reflex events in the flexor reflex is greater than in the monosynaptic reflex, because of the reverberating circuits. EPSPs and IPSPs The arrival of action potentials in the axon terminals of Ia afferents from a muscle is rapidly followed by an EPSP in the motoneurones of that muscle. Summation: Spatial and Temporal The more afferents that are stimulated the larger the size of the EPSPs; this is because EPSPs summate. The more axon terminals excited, the more transmitter is released, and the greater the size of the EPSP recorded. When the EPSPs depolarise the motoneurone membrane to threshold the initial segment if the axon fires and action potential that is conducted along the length of the axon. The summation of EPSPs in the case opposite is due to simultaneous activity in several afferent axons, and this is known as Spatial Summation. Spatial summation also occurs when Ia afferents arising from synergistic muscles arrive on a motoneurone at the same time. Temporal Summation occurs when several EPSPs generated from a single afferent axon occur within a small space of time. If the second and third action potentials arrive before the previous EPSP has returned to baseline, then the second and third EPSPs start when the membrane potential is already depolarised. Temporal summation is therefore associated with the arrival of trains of action potentials within the same nerve endings. EPSPs are voltage changes recorded in the post-synaptic neurone: neurotransmitter opens ligand-gated ion channels for a short period of time, and the potential achieved then declines exponentially because of the capacitance of the cell membrane. When ion channels open during the EPSP the membrane potential moves towards the average equilibrium potential of all the ions that can move, i.e. to which the membrane becomes permeable. In the case of the EPSP the equipbrium potential is around zero; this is due to the movement of sodium and potassium ions simultaneously - this is the result of glutamate release from Ia terminals acting on AMPA receptors. IPSPs are generated when an inhibitory transmitter such as glycine acts on the post-synaptic membrane. Ligand-gated chloride channels open and drive the membrane potential towards to equilibrium potential of chloride (~-70mV). |
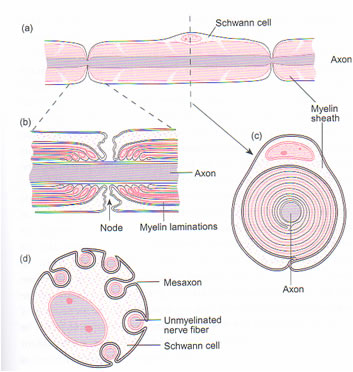
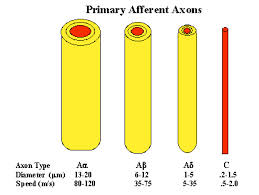
Conduction velocity depends on axonal diameter, and the fastest conduction velocities are in the largest fibres. In contrast the unmyelinated axons have conduction velocities of one hundredth of the fastest myelinated axons.
Interaction of EPSPs and IPSPs EPSPs can summate and depolarise the neuronal membrane; similarly IPSPs can summate and hyperpolarise the membrane. When EPSPs and IPSPs occur at the same time the summate algebraically - i.e. the tend to cancel each other out. It is this balance of excitatory and inhibitory influences that determines whether the neurone fires action potentials, and, if the balance favours excitation, the frequency of action potentials. |
Different reflexes are elicited by different sensory receptors. The physiological classification of sensory receptors depends on the sensory endings and their adequate stimulus. But the conduction velocity of the axons is also a very useful classification as it allows electrophysiologists to identify groups of fibres that are excited by electrical stimuli. Electrical stimulation of different groups of afferent fibres allows the timing of subsequent events to be examined. Consequently there are reflex events associated with stimulation of Ia afferents, and other responses from Ib, Groups II, groups III and IV afferents from muscle nerves. Similarly activation of the A-beta fibres in cutaneous nerves produces no reflex response because these afferents are involved in the sense of touch. Whereas stimulation of A delta and C fibres in skin nerves elicits flexor reflexes, because these afferent axons communicate nociceptive events to the spinal cord. |
|