 * *
Layers of the Cerebellar Cortex
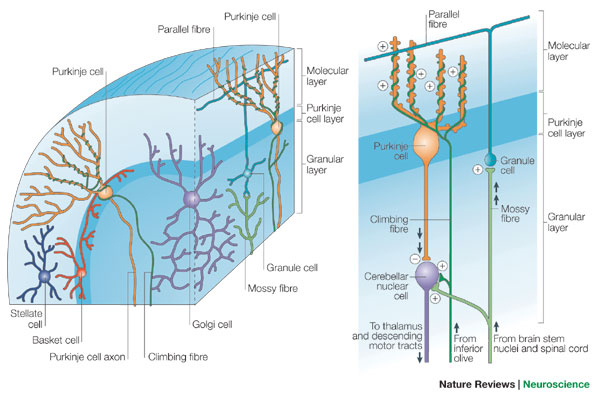
The cerebellar cortex has three layers - a surface (molecular) layer of parallel fibres, consisting of the axons of granule cells and the dendrites of Purkinje cells; the cell bodies of the granule cells are densely packed in the innermost layer - the granular layer; and the Purkinje cell layer, one cell in thickness, between them, and is the main output pathway for the cerebellum.
The axons of granule cells pass towards the surface of the molecular layer and bifurcate, sending their collaterals in opposite directions within the folia where they become parallel fibres, and make excitatory, glutamatergic, synapses on the dendrites of hundreds of Purkinje Cells.
The planar arbors of Purkinje cell dendrites are oriented perpendicular to the parallel fibers, and this arrangement resembles wires running between electricity pylons. Purkinje cells can therefore extract information from the molecular layer, and pass it to the deep cerebellar nuclei, where they release GABA and have inhibitory effects.
Inputs to Purkinje Cells
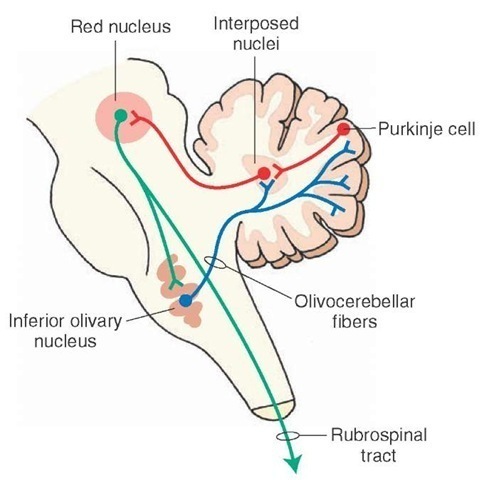
Purkinje cells have two types of input: (a) the parallel fibres of the molecular layer (the axons of granule cells), and (b) climbing fibres which originate in the inferior olive and have a very strong excitatory influence on the Purkinje cells.
Climbing fibres
The name 'climbing fibers' is used because the axons wrap themselves around the Purkinje cell dendrites. Every Purkinje cell receives a very powerful excitatory synaptic input from each climbing fibre, each of which innervates ony a small number of Purkinje cells; this is in contrast to the very large number of weaker excitatory inputs Purkinje cells receive from parallel fibres. Climbing fibres arise from the contralateral inferior olivary nucleus and excite Purkinje cells AND an associated cluster of neurones in the deep cerebellar nuclei.
Mossy Fibres
The parallel fibres of the cerebellar cortex arise from granule cells and are excited by axons originating in the pontine nuclei, that carry a copy of the motor command sent to the motoneurones. These axon terminals on granule cells are called mossy fibres, because of the tufted appearance of their synapses, and they also have axon collaterals that end on neurones of the deep cerebellar nuclei.
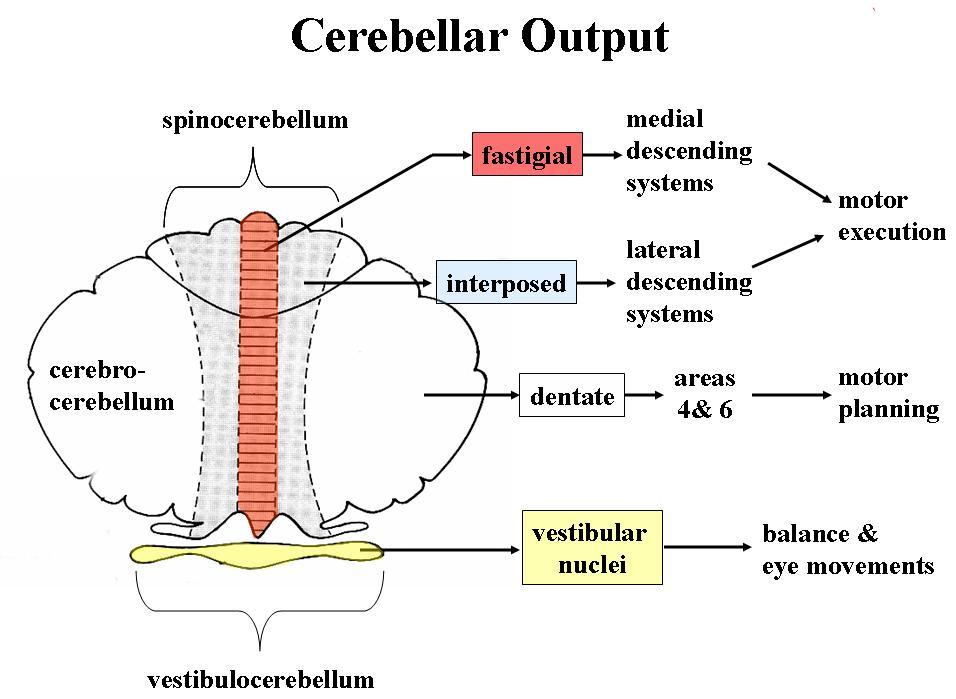
Cerebellar Modules and Cerebellar Cortical Microzones
From a functional point of view, the cerebellum is divided into hundreds of independent modules, all of which have a similar basic structure, but each handles information from different sources, and relays the module output to a different destination.
Each module consists of
- a small group of neurones in the inferior olive that give rise to climbing fibres,
- a spatially discrete group of Purkinje cells and
- a few neurons in a deep cerebellar nucleus on which the Purkinje cells synapse.
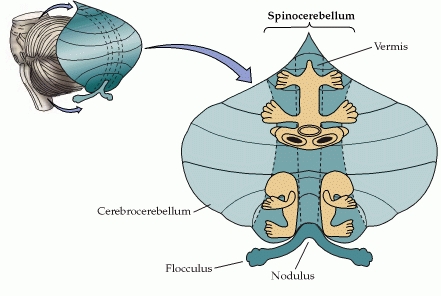
A module consists of about 1000 Purkinje cells that extract information from the millions of parallel fibres that converge on them. All of these Purkinje cells have the same somatotopic receptive field, and occupy a long, continuous narrow strip of cerebellar cortex, aligned perpendicular to the cortical folds. These 1000 Purkinje cells all send their axons to the same small cluster of neurones within the deep cerebellar nuclei. This linear area of cerebellar cortex is called a microzone.
 * *
This diagram shows the arrangement of the Purkinje cell dendrites perpendicular to the parallel fibres, and the microzones in the plane of the Purkinje cell dendrites.
The collaterals of a single climbing fiber make contact with some of the Purkinje cells within a module. As the olivary neurones of the microzone are often coupled by gap junctions, their activity is synchronous, as do the Purkinje cells with which they make contact. Climbing fibers produce occasional activity in Purkinje cells, but because they cause calcium entry into Purkinje cells, they can have longer lasting effects due to the activation of second messenger cascades within the neurone.
These longer-lasting changes can alter the strength of the parallel fibre input to the cell, and hence alter the gain of the system. This may be used as a mechanism for motor learning, as each module becomes more sensitive to its array of inputs that are a marker for a selected motor task.
Divergence and Convergence
While the number of input cells and ouput neurones in the cerebellum may be relatively small, but there is considerable divergence in the input system, resulting in millions of parallel fibres within each module, and considerable convergence in the output system. It can be seen that these modules carry infomation in one direction through the module, without internal feedback; this arrangement is known as a feedforward system.
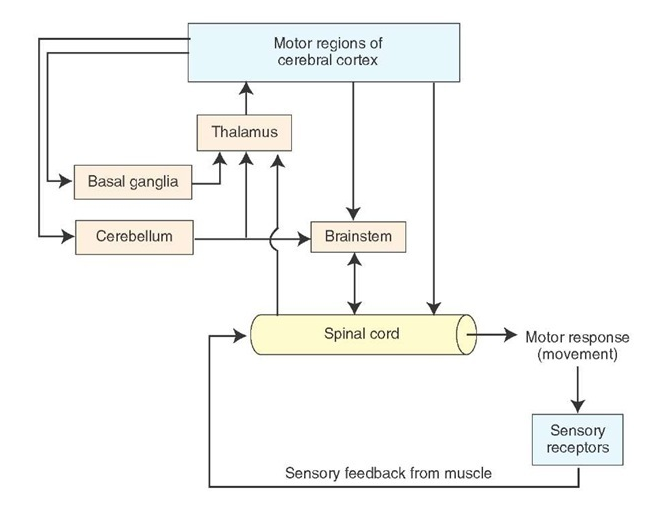
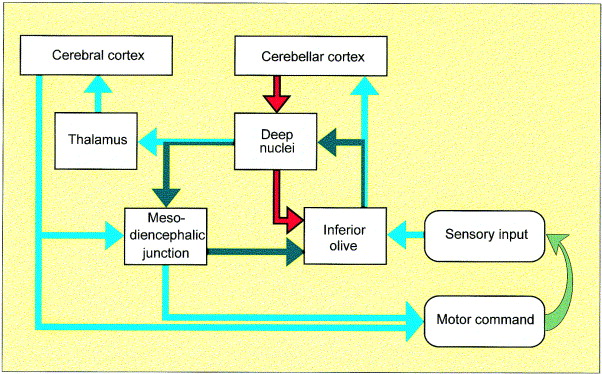
Two-way communication between the cerebellum and the brainstem
The parallel fibres provide the Purkinje cells with a massive amount of of somatosensory, visual and motor information at any moment in time, and this information is changing all the time. The role of the Purkinje cells seems to be to recognise the features of important motor events at any one moment, and the changes that occur moment by moment. The cerebellar modules appear to be able recognise defined combinations of inputs, whch may be reinforce during motor learning.
The importance of the climbing fibres within each cerebellar module raises questions as to what causes them to become active. It is interesting that they have inputs from proprioceptive and cutaneous sensory system and from the cortex and red nucleus; the last of these could potentially carry information about recent processing within the cerebellum. And there are some direct pathways from the deep cerebellar nuclei to discrete areas of the inferior olivary nuclei. So while there is no doubt that the cerebellum itself is a feedforward system, pathways are present in the brainstem that can activate climbing fibres and signal recent computations made within this neural network.
Direct connections from the cerebellum to the brainstem
There are some small but direct connections between the cerebellum and the brainstem, including some that project to the inferior olive and others that project to the vestibular nuclei.
Noradrenergic and Serotoninergic Inputs to the Cerebellum
The locus coeruleus of the brainstem is the origin of a noradrenergic pathway that makes contact with Purkinje cells throughout the cerebellum; this pathway has a neuromodulatory function possibly associated with motor learning.
Serotoninergic pathways also project throughout the entire cerebellum and originate in the rostral raphe nuclei of the brainstem. Serotonin is also involved in modulating the effects of synaptic inputs within the cerebellar cortex.
Motor Learning
The cerebellum is involved in some types of motor learning, particularly when fine adjustments to a motor task are required, such as in playing a violin or piano. Quite how this is accomplished is not fully understood, but the climbing fibre signal has been likened to an error signal by some, and a teaching signal by others; other authors suggest that other parts of the brain are also necessary for learning motor tasks.
Basket Cells and Stellate Cells
Basket cells and Stellate cells are inhibitory interneurons in the molecular layer of the cerebellum. They appear to be able to modulate the activities of Purkinje cells by collecting information from the parallel fibres and inhibiting the Purkinje cells. They may be involved in motor learning.
The distribution of the axons of basket cells are confined largely to a single module and help to ensure that their inhibitory interactions are confined to the microzone to which they belong. |