|
Neuronal Basis of the EEG and its Rhythms Top |
|
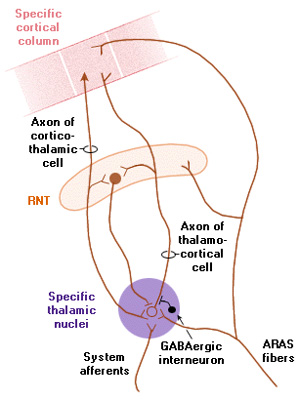
Reverberating Circuits The cortex and thalamic nuclei are connected by excitatory axons in both directions: to and from the cortex. If neurones in a cortical column fire action potentials, they excite their own group of thalamic neurones, which in turn re-excite neurones in the same cortical column. There is plenty of evidence that this type of circuit is active throughout the cortex, and is responsible for the simultaneous rhythmical activity of cortical neurones in the awake, resting state. Modulation of the rhythm of activity in this circuit is achieved by adding another neurone that has its own intrinsic rhythmicity, and its action potentials inhibit the ongoing activity of the reverberating circuit, and determines the ebb and flow of cortical activity. If the inhibitory neurone fires, say, 10 times a second, then the reverberating circuit adopts the rhythmicity imposed on it, i.e. it might have an intrinsic rhythm of 10 Hz. In the discussion below, the looped pathways linking cortical neurones and relay neurones can be inhibited by GABA-ergic neurones in the Reticular Nucleus of the Thalamus, as shown in the illustration below. This rhythmic activity can be disturnbed by sensory or other inputs, as shown. |
|
The Reticular Nucleus of the Thalamus (RTN) forms a 'shell' or a 'skin' around the thalamus, and its neurones have some special characteristics. They are pacemakers because of the nature of their ion channels, and fire action potentials in the absence of external inputs. The also have widespread connections with the specific relay nuclei of the thalamus (for the somatosensory, visual and auditory systems), and because of these projections their activity can affect the whole of the cerebral cortex. They do this by inhibiting the specific relay nuclei using GABA as a neurotransmitter. Cortico-thalamic reverberating circuits are formed by the axons of pyramidal cells that excite both thalamic nuclei - the reticular nucleus and the relay nuclei. So excitation of the cortical pyramidal cells leads to depolarisation of the relay nuclei which in turn re-excite the cortical neurones; the level of activity in this circuit is moderated by the tonic inhibitory activity of the reticular nucleus. In the absence of external sensory inputs the rhythmical activity of the cortico-thalamo-cortical loop is determined by the pacemaker cells of the reticular nucleus of the thalamus, whose projections are widepread in the relay nuclei. So this thalamic circuitry determines how pyramidal cells throughout the cortex fire synchronously in the absence of external sensory inputs. When the ascending reticular activating system becomes active it adds further mild depolarisations at the level of the relay and reticular thalamic nuclei and within the cortex; this input disrupts the existing rhythm and leads to desynchronisation of the EEG. Another special arrangement within this circuit is that the pyramidal neurones of each cortical column project selectively to their own part of the relay and reticular nuclei. Sometimes the cortex and thalamus are said to be 'reciprocally connected' and the circuits are sometimes called 'reverberating circuits'. |
While the reticular nucleus is responsible for synchronising many cortical columns in the absence of external stimuli, excitatory inputs from sensory systems or the ascending reticular activating system (ARAS) alter the basic frequency of each circuit. ARAS produces a mild depolarisation of thalamic and cortical neurones that sensitizes them to the presence of other inputs, as well as desynchronising the EEG. Cholinergic inputs from the nucleus basalis, the Nucleus of Meynert, and the pedunculo-pontine nucleus act through nicotinic receptors in the thalamus and across the cerebral cortex to achieve this desynchronisation. |
| Circuits involved in Synchronisation and Desynchronisation of the EEG Top |
|
At the onset of sleep, activity in ARAS is reduced: the cholinergic and noradrenergic systems become less active, allowing the basic rhythmical activity of the reticular nucleus to be re-established, and the reverberating cortico-thalamo-cortical circuits become synchronously active. At the same time, the relay nuclei of the thalamus receive less depolarisation from the cholinergic system, and are consequently less sensitive to environmental stimuli, as happens in deep sleep. In Stage 2 the EEG shows sleep spindles, an intermediate EEG state between the awake state and deeper sleep. During the spindle there is a short period of activation of the cortex, alongside the general slowing of the EEG. In deeper sleep (stages 3 and 4) the anterior hypothalamus (VLPO) plays a major role in inhibiting the thalamus and cortex, using GABA and galanin. Lastly, during REM sleep, one cause of the desynchronized EEG traces characteristic of this stage is the influence of the cholinergic neurons on thalamic neurones, which prevents the expression of their rhythmic oscillatory activity by the same mechanisms described above with regard to wakefulness. REM sleep and non-REM sleep are rather different states, in terms of muscle tone and metabolism. In deep sleep (Stage 4), metabolic rate decreases and body temperature falls. In contrast, during REM sleep the metabolic rate rises to normal waking levels, despite the reduction in EMG activity. |
|
|
Neuronal Circuitry involved in Switching between non-REM and REM sleep Top |
|
Neuronal activity during REM Sleep One change that occurs at the the initiation of REM sleep is a rapid decrease of activity in noradrenergic and serortoninergic neurones, so much so that they fall silent. This is believed to be the result of inhibition by a group of GABAergic neurons that exist in the upper pons. There are some other groups of brainstem cholinergic neurones become very active during REM sleep, but are inactive when the subject is awake; these neurones are located in an anatomically complex area of the upper brainstem, referred to by some as the parabrachial region, and by others as the oral pontine reticular nucleus. Eye movements are normally coordinated in the adjacent paramedian pontine reticular formation (PPRF), and at the onset of REM sleep, waves of activity can be recorded from the upper pons, the lateral geniculate body and the occipital cortex (sometimes called Ponto-Geniculo-Occipital or PGO waves). So the upper pons seems to be of some importance in initiating the eye movements in general, but also during REM sleep. The pathways that give rise to dreaming are less clear; however some studies have shown that subjects given hypnotic drugs that abolish REM sleep do not dream. Also narcoleptic patients who have an excess of REM sleep also have vivid dreams, even during their short narcoleptic naps - so the association between REM sleep and dreaming is well established. The neurological nature of the link between REM sleep and dreaming is uncertain, but some authors believe dreaming is accompanied by activity in the frontal and visual cortices and the hippocampus. Another feature of REM sleep is the absence of EMG activity, causing generalised muscular relaxation. This hypotonic state appears to depend on descending pathways the excite the glycine-containing inhibitory interneurones in the ventral horn of the spinal cord. The switching mechanisms that result in this state are complex and not entirely clear, but the brainstem appears to be able to coordinated the separate components of REM sleep. It should be remembered that the role of serotonin is complicated because this amine helps to induce sleep by exciting VLPO neurones in the hypothalamus. Arousal Some neurons are completely silent during REM sleep, but become active at the time of awakening; the tubero-mamillary neurones are one example of this behaviour, and these release histamine at the terminations throughout the CNS. |
|