A. The Utricle and the Saccule:
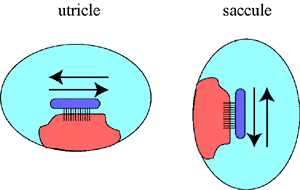
1. The Utricle and the Saccule: The utricle and the saccule are fluid-filled spaces. They contain a macula which is oriented in the horizontal direction ( = utricle) or in the vertical direction (= saccule). These two organs therefore detect movements in the horizontal or in the vertical direction. |
|
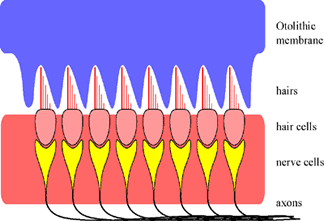
2. The Macula: The macula is the sensor in these two organs. It essentially consists of a row of hair cells. The hairs are connected to a gelatinous mass called the otholithic membrane. The other side of the hair cell is connected to a nerve. This is the beginning of the vestibular nerve. |
|
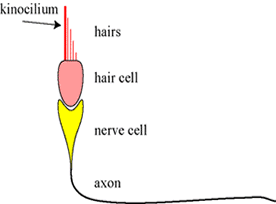
3. The Hair Cell: In contrast to the hair cells in the cochlea, there is also an orientation in the configuration of the hairs in the macula. One of the hairs is longer and thicker than the others. This special hair is called a kinocilium (plural kinocilia) and is, as it where, a point of reference for these hairs. |
|
4. How does it work? When the head starts to move in one direction, the otolithic membrane will move in the opposite direction. This is due to the inertia of the otolith. Because the hair cells move in one direction and the otolithic membrane in the other, all the hairs will bend. As we have seen before in the organ of Corti, this will influence stretch channels in the neighborhood to open, thereby depolarizing the membrane and generating action potentials. |
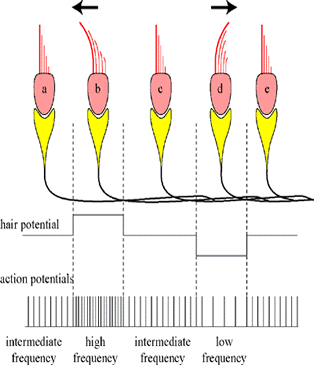 |
5. Forward and backward or up-and-down? But in the saccule and the utricle, there is an additional problem. These two organs must also detect whether the horizontal movement is forward or backward (or in the case of a vertical movement up or down). As shown in the diagram, this is solved in the following manner. If the hairs bend towards the kinocilium, then there will be a depolarization in the hair cell. If the hairs bend the other way, away from the kinocilium, then the hair cell will hyperpolarize. |
|
6. Frequency modulation: The final step is to transduce (=translate) the depolarization into action potentials. As in the organ of Corti, depolarization causes more action potentials and the frequency becomes higher. This is because the polarity moves closer to the threshold. The opposite happens when the polarity moves away from the threshold. Then, during hyperpolarization, there will be less action potentials; the frequency is lower. This transduction from amplitude to frequency is (technically) called "frequency modulation" (like in the radio! FM vs. AM). |
|
7. Acceleration versus velocity. It is important to realize that it is the acceleration that stimulates the otolithic membrane to shift, thereby bending the hairs, not the velocity itself. If it had been the velocity that shifted the otoliths, then we would never be able to drive in a car or fly in a plane. No, it is the change in velocity (=acceleration or deceleration) that shifts the otoliths and that is what we feel. |
8. Frequency modulation. Note that when the utricle or the saccule are in rest, that then the hair cells still generate action potentials. In other words, it is actually telling the brain that it is working and that there is no acceleration or deceleration. But if the frequency increases or decreases, then it can report how much (the change in frequency) and in which direction (accelerating or decelerating). |